The Meselson-Stahl Experiment (1957–1958), by Matthew Meselson and Franklin Stahl
 Editor's note: Madeleine Howell-Moroney created the above image for this article. You can find the full image and all relevant information here.
Editor's note: Madeleine Howell-Moroney created the above image for this article. You can find the full image and all relevant information here.
In an experiment later named for them, Matthew Stanley Meselson and Franklin William Stahl in the US demonstrated during the 1950s the semi-conservative replication of DNA, such that each daughter DNA molecule contains one new daughter subunit and one subunit conserved from the parental DNA molecule. The researchers conducted the experiment at California Institute of Technology (Caltech) in Pasadena, California, from October 1957 to January 1958. The experiment verified James Watson and Francis Crick´s model for the structure of DNA, which represented DNA as two helical strands wound together in a double helix that replicated semi-conservatively. The Watson-Crick Model for DNA later became the universally accepted DNA model. The Meselson-Stahl experiment enabled researchers to explain how DNA replicates, thereby providing a physical basis for the genetic phenomena of heredity and diseases.
The Meselson-Stahl experiment stemmed from a debate in the 1950s among scientists about how DNA replicated, or copied, itself. The debate began when James Watson and Francis Crick at the University of Cambridge in Cambridge, England, published a paper on the genetic implications of their proposed structure of DNA in May 1953. The Watson-Crick model represented DNA as two helical strands, each its own molecule, wound tightly together in a double helix. The scientists claimed that the two strands were complementary, which meant certain components of one strand matched with certain components of the other strand in the double helix.
With that model of DNA, scientists aimed to explain how organisms preserved and transferred the genetic information of DNA to their offspring. Watson and Crick suggested a method of self-replication for the movement of genetic information, later termed semi-conservative replication, in which DNA strands unwound and separated, so that each strand could serve as a template for a newly replicated strand. According to Watson and Crick, after DNA replicated itself, each new double helix contained one parent strand and one new daughter strand of DNA, thereby conserving one strand of the original double helix. While Watson and Crick proposed the semi-conservative model in 1953, the Meselson-Stahl experiment confirmed the model in 1957.
In 1954, Max Delbrück at Caltech published a paper that challenged the Watson-Crick Model for DNA replication. In his paper, Delbrück argued that the replication process suggested by Watson and Crick was unlikely because of the difficulty associated with unwinding the tightly-wound DNA structure. As an alternative, Delbrück proposed that instead of the entire structure breaking apart or unwinding, small segments of DNA broke from the parent helix. New DNA, Delbrück claimed, formed using the small segments as templates, and the segments then rejoined to form a new hybrid double helix, with parent and daughter segments interspersed throughout the structure.
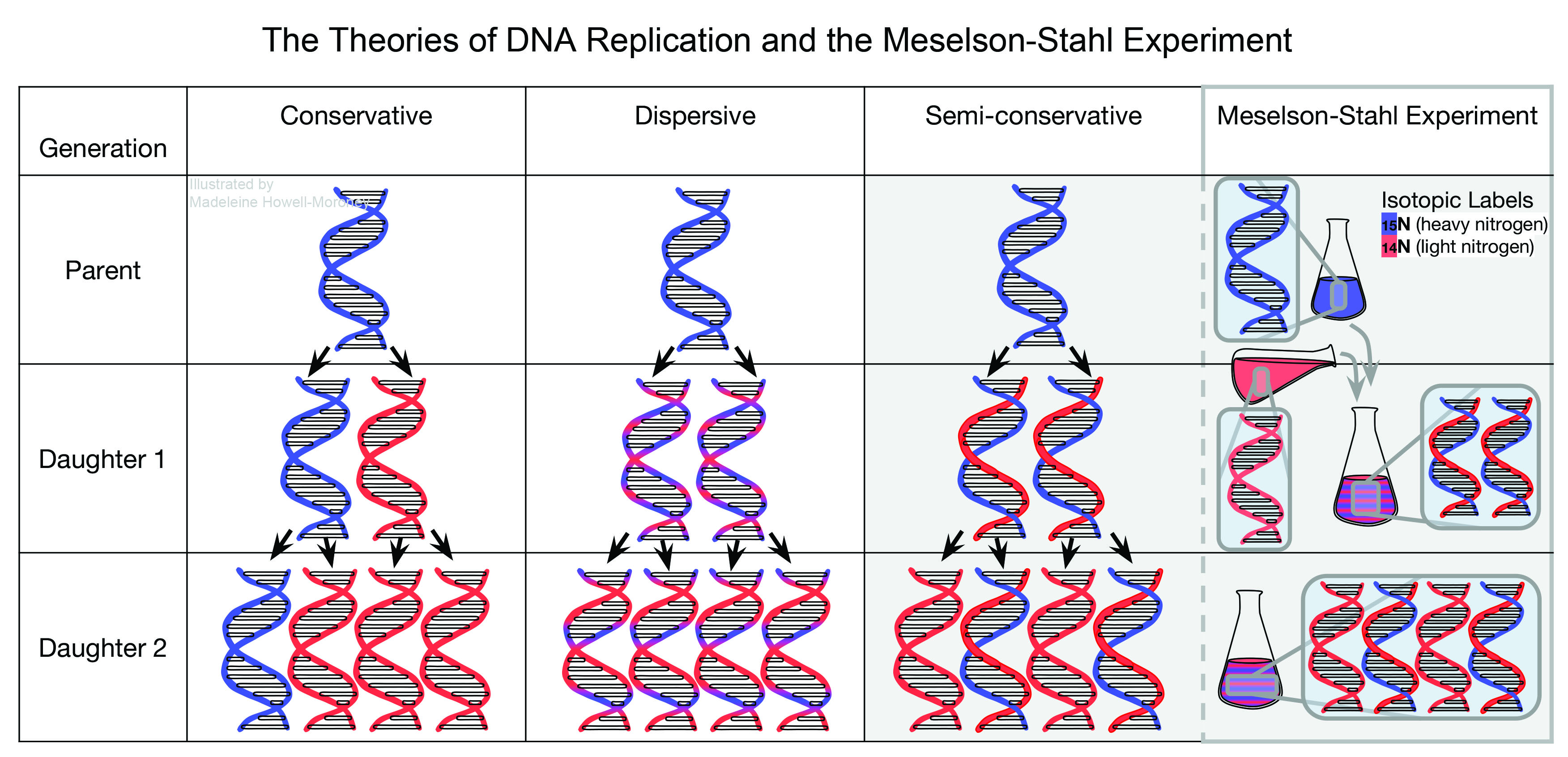
After the release of Delbrück´s paper, many scientists sought to determine experimentally the mechanism of DNA replication, which yielded a variety of theories on the subject by 1956. Delbrück and Gunther Stent, a professor at the University of California, Berkeley, in Berkeley, California, presented a paper in June 1956 at a symposium at Johns Hopkins University in Baltimore, Maryland, which named and summarized the three prevailing theories regarding DNA replication at the time: semi-conservative, dispersive, and conservative. Delbrück and Stent defined conservative replication as a replication mechanism in which a completely new double helix replicated from the parent helix, with no part of the parent double helix incorporated into the daughter double helix. They described the semi-conservative process as Watson and Crick suggested, with half of the parental DNA molecule conserved in the daughter molecule. Lastly, Delbrück and Stent summarized Delbrück´s dispersive model, in which parental DNA segments distribute throughout the daughter DNA molecule. Delbrück and Stent´s paper provided the background for the Meselson-Stahl experiment.
In 1954, prior to publication of Delbrück´s initial challenge of the Watson-Crick model, Matthew Meselson and Franklin Stahl had joined the DNA replication discussion. During the spring of 1954, Meselson, a graduate student studying chemistry at Caltech, visited Delbrück´s office to discuss DNA replication. According to historian of science Frederic Holmes, during that meeting Meselson began brainstorming ways to determine how DNA replicated. In the summer of 1954, Meselson met Stahl at the Marine Biological Laboratory in Woods Hole, Massachusetts. Stahl, a graduate student studying biology at the University of Rochester in Rochester, New York, agreed to study DNA replication with Meselson the following year at Caltech.
Meselson and Stahl began their collaboration in late 1956. By that time, Stahl had completed his PhD and Meselson had completed the experiments for his PhD, which he received in 1957. They worked on a variety of projects, including DNA replication. All of their projects, however, involved a method first devised by Meselson in 1954, called density-gradient centrifugation. Density-gradient centrifugation separates molecules based on their densities, which depend on the molecular weights of the molecules.
Meselson and Stahl used density-gradient centrifugation to separate different molecules in a solution, a method they later used to separate DNA molecules in a solution. In density gradient centrifugation, a solution is placed in an ultracentrifuge, a machine that spins the samples very fast on the order of 140,000 times the force of gravity or 44,770 revolutions per minute (rpm). As the samples spin, denser substances are pushed toward the bottom, while less dense substances distribute according to their weight in the centrifuge tube. By the end of centrifugation, the molecules reach a position called equilibrium, in which the molecules stop moving and remain in a gradient. The position of the molecules at equilibrium is dependent on the density of the molecule. Meselson and Stahl measured the areas in which DNA was at the highest concentration. Higher concentrations were represented by darker bands of DNA in the centrifuged sample. Stahl represented those bands on a graph, so that the peaks represented locations in the gradient where there was the highest concentration of molecules. Multiple peaks meant that molecules of different densities separated out of the solution.
To describe how DNA replicated, Meselson and Stahl needed to distinguish between parental and daughter DNA. They achieved that by modifying the molecules so each kind had a different density. Then Meselson and Stahl could separate the molecules using density-gradient centrifugation and analyze how much parental DNA was in the new daughter helices after every replication cycle. First they tried to alter the density of parental DNA by substituting a one nucleotide base, thymidine, with a heaver but similar DNA nucleotide base, 5-bromouriacil (5-BU). However, Meselson and Stahl struggled to substitute enough units of 5-BU into the DNA molecules to make the parental DNA significantly denser than normal DNA.
By July 1957, Meselson and Stahl successfully incorporated the heavy substitution in parental DNA, but the type of DNA they used still caused problems. Meselson and Stahl first used DNA from a specific type of virus that infects bacteria, called a bacteriophage. However, bacteriophage DNA not only broke apart in solution during centrifugation, but also replicated too quickly for the distribution of DNA to be adequately measured after each cycle. Consequently, Meselson and Stahl struggled to see clear locations within the density gradient with the highest concentration of bacterial DNA. Therefore, in September 1957, Meselson and Stahl switched to using the DNA from the bacteria Escherichia coli (E. coli). E. coli DNA formed clearer concentration peaks during density gradient centrifugation.
At around the same time, in addition to changing the source DNA, Meselson and Stahl also changed the type of density label they used, from substitution labels to isotope labels. An isotope of an element is an atom with the same number of positive charged nuclear particles or protons, and a different number of uncharged particles, called neutrons. A difference in neutrons, for the most part, does not affect the chemical properties of the atom, but it alters the weight of the atom, thereby altering the density. Meselson and Stahl incorporated non-radioactive isotopes of nitrogen with different weights into the DNA of E. coli. As DNA contains a large amount of nitrogen, so long as the bacteria grew in a medium containing nitrogen of a specified isotope, the bacteria would use that nitrogen to build DNA. Therefore, depending on the medium in which E. coli grew, daughter strands of newly replicated DNA would vary by weight, and could be separated by density-gradient centrifugation.
Starting in October 1957, Meselson and Stahl conducted what later researches called the Meselson-Stahl experiment. They grew E. coli in a medium containing only the heavy isotope of nitrogen (15N) to give the parental DNA a higher than normal density. As bacteria grow, they duplicate, thereby replicating their DNA in the process. The researchers then added an excess of light isotopes of nitrogen (14N) to the heavy nitrogen environment.
Meselson and Stahl grew E. coli in the 14N isotope environment for all subsequent bacterial generations, so that any new DNA strands produced were of a lower density than the original parent DNA. Before adding 14N nitrogen, and for intervals of several bacterial generations after adding light nitrogen, Meselson and Stahl pulled samples of E. coli out of the growth medium for testing. They centrifuged each sample for initial separation, and then they added salt to the bacteria so that the bacteria released its DNA contents, allowing Meselson and Stahl to analyze the samples.
Next, Meselson and Stahl conducted density gradient centrifugation for each DNA sample to see how the parental and daughter DNA distributed according to their densities over multiple replications. They added a small amount of each sample of bacterial DNA to a cesium chloride solution, which when centrifuged had densities within the range of the bacterial DNA densities so that the DNA separated by density. The researchers centrifuged the DNA in an ultracentrifuge for twenth hours until the DNA reached equilibrium. Using ultraviolet light (UV), the researchers photographed the resulting DNA bands, which represented peaks of DNA concentrations at different densities. The density of the DNA depended on the amount of 15N or 14N nitrogen present. The more 15N nitrogen atoms present, the denser the DNA.
For the bacterial DNA collected before Meselson and Stahl added 14N nitrogen, the UV photographs showed only one band for DNA with 15N nitrogen isotopes. That result occurred because the DNA from the first sample grew in an environment with only 15N nitrogen isotopes. For samples pulled during the first replication cycle, the UV photographs showed fainter the 15N DNA bands, and a new DNA band formed, which represented half 15N DNA nitrogen isotopes and half 14N DNA nitrogen isotopes. By the end of the first replication cycle, the heavy DNA band disappeared, and only a dark half 15N and half 14N DNA band remained. The half 15N half 14N DNA contained one subunit of 15N nitrogen DNA and one subunit of 14N nitrogen DNA. The data from the first replication cycle indicated some distribution of parental DNA, therefore ruled out conservative replication, because only parental DNA contained 15N nitrogen isotopes and only parental DNA could represent the 15N nitrogen isotopes in daughter DNA.
The same trends continued in future DNA replication cycles. As the bacteria continued to replicate and the bacterial DNA replicated, UV photographs showed that the band representing half 15N half 14N DNA depleted. A new band, representing DNA containing only 14N nitrogen isotopes or light DNA, became the prevalent DNA band in the sample. The depletion of the half 15N half 14N band occurred because Meselson and Stahl never re-introduced 15N nitrogen, so the relative amount of 15N nitrogen DNA decreased. Meselson and Stahl then mixed the samples pulled from different replication cycles and centrifuged them together. The UV photograph from that run showed three bands of DNA with the half 15N half 14N DNA band at the midpoint between the 15N DNA band and 14N DNA band, making it an intermediate band. The result indicated that the half 15N half 14N DNA band had a density exactly between the 15N and 14N nitrogen DNA, showing that the DNA in the central band contained half of the 15N nitrogen and half of the 14N nitrogen isotopes, just as predicted by the Watson and Crick model. The exact split between heavy and light nitrogen characterized semi-conservative DNA replication.
Meselson and Stahl made three conclusions based on their results. First, they concluded that the nitrogen in each DNA molecule divided evenly between the two subunits of DNA, and that the subunits stayed intact throughout the observed replication cycles. Meselson and Stahl made that conclusion because the intermediate band had a density halfway between the heavy and light DNA bands. That conclusion made by Meselson and Stahl challenged the dispersive mechanism suggested by Delbrück, which involved breaking the DNA subunits into smaller pieces.
Meselson´s and Stahl´s second conclusion stated that each new DNA double helix contained one parental subunit, which supported semi-conservative replication. Assuming that DNA consists of two subunits, if a parent passes on one subunit of DNA to its offspring, then half of the parental DNA is conserved in the offspring DNA, and half of the parental DNA is not. The researchers made that conclusion because if parental DNA did not replicate in that way, then after the first replication, some DNA double helices would have contained only parental heavy nitrogen subunits or only daughter light nitrogen subunits. That type of replication would have indicated that that some parental DNA subunits did not separate in the semi-conservative fashion, and instead would have supported conservative replication. The presence of one parental subunit for each daughter DNA double helix supported semi-conservative replication.
The third conclusion made by Meselson and Stahl stated that for every parental DNA molecule, two new molecules were made. Therefore, the amount of DNA after each replication increased by a factor of two. Meselson and Stahl related their findings to the structure of DNA and replication mechanism proposed by Watson and Crick.
Before Meselson and Stahl published their findings, word of the Meselson-Stahl results spread throughout Caltech and the scientific community. According to Holmes, Delbrück, who had strongly opposed the semi-conservative method of DNA replication, immediately accepted DNA replication as semi-conservative after seeing the results from the Meselson-Stahl experiment. Some experiments earlier that year had pointed towards semi-conservative replication, and the Meselson-Stahl experiment served to further support semi-conservative replication.
Despite the positive reception of the Meselson-Stahl experiment, years passed before scientists fully accepted the Watson-Crick Model for DNA based on the findings from the Meselson-Stahl experiment. The Meselson-Stahl experiment did not clearly identify the exact subunits that replicated in DNA. In the Watson and Crick model, DNA consisted of two one-stranded DNA subunits, but the Meselson-Stahl experiment also supported models of DNA as having more than two strands. In 1959, Liebe Cavalieri, a scientist at the Sloan-Kettering Institute for Cancer research in New York City, New York, and his research team had produced evidence supporting the theory that DNA consisted of two two-stranded subunits, making DNA a quadruple helix. Cavalieri´s proposal did not contradict the Meselson-Stahl experiment, because the Meselson-Stahl experiment did not define DNA subunits. However, later experiments performed by Meselson on bacteriophage DNA from 1959 to 1961, and experiments performed by John Cairns on E. coli DNA in 1962, settled the debate and showed that each subunit of DNA was a single strand.
As described by Holmes, many scientists highly regarded the Meselson-Stahl experiment. Scientists including John Cairns, Gunther Stent, and James Watson all described the experiment as beautiful in both its performance and simplicity. Holmes also described the academic paper published by Meselson and Stahl on their experiment as beautiful because of its concise descriptions, diagrams, and conclusions. The Meselson-Stahl experiment appeared in textbooks decades after Meselson and Stahl performed the experiment. In 2001, Holmes published Meselson, Stahl, and the Replication of DNA: A History of "The Most Beautiful Experiment in Biology," which told the history of the experiment.
The Meselson-Stahl experiment gave a physical explanation for the genetic observations made before it. According to Holmes, for scientists who already believed that DNA replicated semi-conservatively, the Meselson-Stahl experiment provided concrete evidence for that theory. Holmes stated that, for scientists who contested semi-conservative replication as proposed by Watson and Crick, the Meselson-Stahl experiment eventually changed their opinions. Either way, the experiment helped scientists´ explain inheritance by showing how DNA conserves genetic information throughout successive DNA replication cycles as a cell grows, develops, and reproduces.
Sources
- Cairns, John. "A Minimum Estimate for the Length of the DNA of Escherichia coli Obtained by Autoradiography." Journal of Molecular Biology 4 (1962): 407–9.
- Cavalieri, Liebe F., Barbara Hatch Rosenberg, and Joan F. Deutsch. "The Subunit of Deoxyribonucleic Acid." Biochemical and Biophysical Research Communications 1 (1959): 124–8.
- Davis, Tinsley H. "Meselson and Stahl: The Art of DNA Replication." Proceedings of the National Academy of Sciences 101 (2004): 17895–6. http://www.pnas.org/content/101/52/17895.long (Accessed April 18, 2017).
- Delbrück, Max. "On the Replication of Deoxyribonucleic Acid (DNA)." Proceedings of the National Academy of Sciences 40 (1954): 783–8. http://www.pnas.org/content/40/9/783.short (Accessed April 18, 2017).
- Delbrück, Max and Gunther S. Stent. "On the Mechanism of DNA Replication." In McCollum-Pratt Symposium on the Chemical Basis of Heredity, eds. William D. McElroy and Bentley Glass, 699–736. Baltimore: Johns Hopkins University Press, 1956.
- Holmes, Frederic L. Meselson, Stahl, and the Replication of DNA: a History of "The Most Beautiful Experiment in Biology." New Haven: Yale University Press, 2001.
- "Interview with Matthew Meselson." Bioessays 25 (2003): 1236–46.
- Judson, Horace Freeland. The Eighth Day of Creation: Makers of the Revolution in Biology. Cold Spring Harbor: Cold Spring Harbor Laboratory Press, 1996.
- Levinthal, Cyrus. "The Mechanism of DNA Replication and Genetic Recombination in Phage." Proceedings of the National Academy of Sciences 42 (1956): 394–404. http://www.pnas.org/content/42/7/394.short (Accessed April 18, 2017).
- Litman, Rose M. and Arthur B. Pardee. "Production of Bacteriophage Mutants by a Disturbance of Deoxyribonucleic Acid Metabolism." Nature 178 (1956): 529–31.
- Meselson, Matthew. "The Semi-Conservative Replication of DNA." iBioMagazine 5 (2011). https://www.ibiology.org/ibiomagazine/issue-5/matthew-meselson-the-semi-conservative-replication-of-dna.html (Accessed April 18, 2017).
- Meselson, Matthew, and Franklin W. Stahl. "The Replication of DNA in Escherichia Coli." Proceedings of the National Academy of Sciences 44 (1958): 671–82. http://www.pnas.org/content/44/7/671.long (Accessed April 18, 2017).
- Meselson, Matthew, and Jean Weigle. "Chromosome Breakage Accompanying Genetic Recombination in Bacteriophage." Proceedings of the National Academy of Sciences 47 (1961): 857–68. http://www.pnas.org/content/47/6/857.short (Accessed April 18, 2017).
- Meselson, Matthew, Franklin W. Stahl, and Jerome Vinograd. "Equilibrium Sedimentation of Macromolecules in Density Gradients." Proceedings of the National Academy of Sciences 43 (1957): 581–8. http://www.pnas.org/content/43/7/581.short (Accessed April 18, 2017).
- Taylor, J. Herbert, Philip S. Woods, and Walter L. Hughes. "The Organization and Duplication of Chromosomes as Revealed by Autoradiographic Studies Using Tritium-Labeled Thymidine." Proceedings of the National Academy of Sciences 43 (1957): 122–8. http://www.pnas.org/content/43/1/122.short (Accessed April 18, 2017).
- Watson, James D., and Francis H C Crick. "Molecular Structure of Nucleic Acids: A Structure for Deoxyribose Nucleic Acid." Nature 171 (1953): 737–8. https://profiles.nlm.nih.gov/ps/access/SCBBYW.pdf (Accessed April 18, 2017).
- Watson, James D., and Francis H C Crick. "Genetical Implications of the Structure of Deoxyribonucleic Acid." Nature 171 (1953): 964–7. https://profiles.nlm.nih.gov/ps/access/SCBBYX.pdf (Accessed April 18, 2017).
- Weigle, Jean, and Matthew Meselson. "Density Alterations Associated with Transducing Ability in the Bacteriophage Lambda." Journal of Molecular Biology 1 (1959): 379–86.
Keywords
Editor
How to cite
Publisher
Handle
Rights
Articles Rights and Graphics
Copyright Arizona Board of Regents Licensed as Creative Commons Attribution-NonCommercial-Share Alike 3.0 Unported (CC BY-NC-SA 3.0)